Рептилии и млекопитающие, Животные: млекопитающие, рептилии, беспозвоночные, амфибии, птицы

Они населяют все континенты. Wood, Jr. What This Природа. Без ранга:.

Среди вымерших древних рептилий известны высокоподвижные формы такие как архозавры: наземные крокодиломорфы , динозавры и птерозавры , а также лепидозавры : мозазавры , ихтиозавры и завроптеригии , строение тела и предполагаемый образ жизни которых говорят о способности поддерживать продолжительную мышечную активность без утомления, что соответствует более интенсивному, чем у современных пресмыкающихся, обмену веществ, и работе мышц преимущественно по аэробному механизму, требующему интенсивного снабжения кислородом.
Это в свою очередь должно было сопровождаться совершенно другой физиологией организма полное разделение желудочков сердца, повышение эффективности дыхания за счёт отделения воздухоносных путей от ротовой полости, появление эффективных механизмов терморегуляции и так далее. Данные изменения отчасти прослеживаются на палеонтологическом материале в линии рептилий, ведущей к птицам.
В настоящее время надёжно показано существование теплокровности как минимум у некоторых групп динозавров и предков современных крокодилов, а также плезиозавров , мозазавров и ихтиозавров [64] [65] [68]. Как и у большинства хордовых животных, центральная нервная система пресмыкающихся представлена головным из 5 отделов и спинным мозгом.
Головной мозг расположен внутри черепа. Ряд важных особенностей отличает головной мозг пресмыкающихся от головного мозга земноводных. Нередко говорят [ кто? От головного мозга отходят 12 пар черепных нервов.
В спинном мозге разделение на белое и серое вещество более отчётливо, чем у амфибий. От спинного мозга отходят сегментальные спинномозговые нервы, образуя типичное плечевое и тазовое сплетение. Отчётливо выражена вегетативная нервная система симпатическая и парасимпатическая в виде цепи парных нервных ганглиев. Пресмыкающиеся имеют шесть основных органов чувств , из которых зрение имеет решающее значение. Для пресмыкающихся характерно дыхание всасывающего типа путём расширения и сужения грудной клетки при помощи межрёберной и брюшной мускулатуры.
Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи , ведущие в лёгкие. Как и у земноводных, лёгкие пресмыкающихся имеют мешкообразное строение, хотя их внутренняя структура намного сложнее. Внутренние стенки лёгочных мешков имеют складчатое ячеистое строение, что значительно увеличивает дыхательную поверхность.
Строение лёгких значительно отличается у разных видов и может представлять слабодифференциированные органы с малым количеством ячей, у примитивных видов, таких как гаттерии , среднеразвитые органы с ярко выраженной внутренней структурой, как у большинства чешуйчатых, так и высокоразвитые лёгкие, имеющие почти губчатое строение как у птиц и млекопитающих , у таких рептилий, как вараны, тегу, крокодилы, многие черепахи и крупные змеи.
Водные черепахи имеют более проницаемую кожу, а некоторые виды даже изменили свою клоаку, чтобы увеличить площадь газообмена. Даже с этими адаптациями дыхание никогда полностью не осуществляется без лёгких [71]. Смена воздуха в лёгких у каждой основной группы рептилий осуществляется по-разному. У чешуйчатых лёгкие вентилируются почти исключительно осевой мускулатурой.
Это та же самая мускулатура, которая используется при передвижении. Из-за этого ограничения большинство чешуйчатых вынуждены задерживать дыхание во время интенсивных пробежек.
Некоторые, однако, нашли способ обойти это. Вараниды и некоторые другие виды ящериц используют буккальное сцеживание как дополнение к их обычному «аксиальному дыханию».
Это позволяет животным полностью наполнять свои лёгкие во время бега и, таким образом, длительное время оставаться аэробно активными. Известно, что у ящериц тегу имеется протодиафрагма , отделяющая лёгочную полость от висцеральной.
Хотя на самом деле он не способен к движению, он позволяет сильнее раздувать лёгкие, снимая вес внутренних органов с них [72]. Поскольку тело покрыто чешуйками, кожное дыхание у пресмыкающихся отсутствует исключение составляют мягкотелые черепахи и морские змеи , и лёгкие являются единственным дыхательным органом. У видов, живущих в воде, могут появляться специализированные адаптации, для того чтобы обходиться дольше без атмосферного воздуха, например у морских змей это слизистая выстилка рта, а у черепах это и полость рта и специальные анальные мешки, способные усваивать кислород из воды.
Крокодилы на самом деле имеют мышечную диафрагму, аналогичную диафрагме млекопитающих. Разница в том, что мышцы крокодиловой диафрагмы тянут лобковую кость часть таза, подвижную у крокодилов назад, что опускает печень, тем самым освобождая пространство для расширения лёгких.
Этот тип диафрагмальной установки называют «печёночным поршнем». Дыхательные пути образуют ряд двойных трубчатых камер в каждом лёгком. При вдохе и выдохе воздух проходит через дыхательные пути в одном направлении, создавая таким образом однонаправленный поток воздуха через лёгкие. Аналогичная система обнаружена у птиц, варанов и игуан [73] [74]. У большинства рептилий нет вторичного неба, а это означает, что они должны задерживать дыхание при глотании.
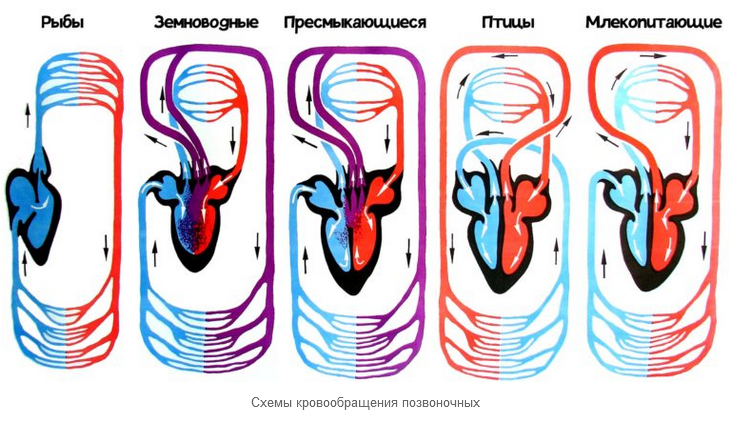
Крокодилы развили костлявое вторичное нёбо, которое позволяет им продолжать дышать, оставаясь под водой и защищать свой мозг от повреждений, связанных с борьбой с добычей. Сцинки семейство Scincidae также в той или иной степени развили костное вторичное небо. Змеи использовали другой подход и вместо этого расширили свои трахеи. Их расширение трахеи торчит изо рта в виде трубки, и позволяет этим животным проглатывать крупную добычу, не страдая от удушья [75]. Как и амфибии, большинство пресмыкающихся обладают трёхкамерным сердцем , состоящим из желудочка и двух предсердий.

Желудочек разделён неполной перегородкой на две половины: верхнюю и нижнюю, и может быть условно разделён на три отдела. Однако трёхкамерное сердце рептилий развилось независимо от такового у земноводных, и различается в плане строения и иногда — функциональности у разных современных групп [76] [77]. Все лепидозавры и черепахи имеют трёхкамерное сердце, состоящее из двух предсердий, одного желудочка с переменными перегородками и двух аорт, ведущих в большой круг кровообращения.
Степень смешения оксигенированной и деоксигенированной крови в трёхкамерном сердце различна в зависимости от вида и физиологического состояния. Было высказано предположение, что это изменение кровотока обеспечивает более эффективную терморегуляцию и более длительное время погружения для водных видов, но не было доказано, что оно является преимуществом и для наземных видов [78]. Будучи архозаврами, крокодилы обладают четырёхкамерным сердцем [64]. Перегородка полностью разделяет желудочек на две половины: правую — венозную и левую — артериальную, таким образом обосабливая полноценное четырёхкамерное сердце, как у млекопитающих и птиц.
Лишь после образования полных перегородок у крокодилов образуется отверстие Панница в ходе эмбриогенеза, допускающее контролируемое смешивание артериальной и венозной крови, что связано со вторичным понижением уровня обмена веществ у этой группы животных [64].
Крокодилы имеют анатомически четырёхкамерное сердце, похожее на птичье, но также имеют две аорты [79]. Например, сердце игуаны, как и большинство чешуйчатых сердец, состоит из трёх камер с двумя аортами и одним желудочком, сердечными непроизвольными мышцами. Основными структурами сердца являются венозный синус, водитель ритма, левое предсердие, правое предсердие, атриовентрикулярный клапан, венозная полость, артериальная полость, лёгочная полость, мышечный гребень, вентрикулярный гребень, лёгочные вены и парные дуги аорты [80].
Некоторые виды чешуйчатых например, питоны и вараны имеют трёхкамерное сердце, которое при сокращении функционально становится четырёхкамерным. Это стало возможным благодаря мышечному гребню, который разделяет желудочек во время диастолы желудочков и полностью разделяет его во время систолы желудочков. Из-за этого гребня некоторые из этих чешуек способны создавать перепады давления в желудочках, эквивалентные тем, которые наблюдаются в сердцах млекопитающих и птиц [81].
Полного разделения на два независимых круга кровообращения у пресмыкающихся не происходит, поскольку венозная и артериальная кровь смешиваются в спинной аорте. Подобно рыбам и амфибиям, подавляющее большинство современных пресмыкающихся — холоднокровные животные. Несмотря на это, многие пресмыкающиеся могут регулировать температуру, перемещаясь из тени на солнце и обратно или меняя цвет, становясь темнее, чтобы согреться, или светлее, чтобы остыть. Сравнительно недавно у ящериц тегу была обнаружена факультативная эндотермия — возможность повышать температуру тела во время брачного периода за счёт эндотермических химических реакций [82].
Вараны в разгар пищевой активности также могут повышать температуру тела за счёт внутренних процессов [83]. Кожистая черепаха, в свою очередь, имеет изолирующую жировую прослойку и за счёт интенсивной мышечной активности может быть примерно на 18 градусов теплее окружающей её воды [84].
Современные нептичьи рептилии холоднокровны, так что они имеют ограниченные физиологические средства поддержания постоянной температуры тела и часто полагаются на внешние источники тепла. Из-за менее стабильной внутренней температуры, чем у птиц и млекопитающих, биохимия рептилий требует ферментов, способных сохранять эффективность в более широком диапазоне температур, чем в случае теплокровных.
В то время как оптимальная температура часто наблюдается, когда животное активно, низкий основной обмен приводит к быстрому падению температуры тела, когда животное неактивно [85] [86]. Как и у всех животных, мышечная деятельность рептилий производит тепло. У крупных рептилий, таких как кожистые черепахи, низкое отношение поверхности к объёму позволяет этому метаболически вырабатываемому теплу держать животных теплее, чем их окружающая среда, даже если у них нет теплокровного метаболизма.
Эта форма гомеотермии называется гигантотермией; было высказано предположение, что она была обычным явлением у крупных динозавров и других вымерших крупных рептилий [87].
Преимущество низкого метаболизма в состоянии покоя заключается в том, что для поддержания функций организма требуется гораздо меньше пищи. Используя колебания температуры окружающей среды или оставаясь холодными, когда им не нужно двигаться, рептилии могут экономить значительное количество энергии по сравнению с эндотермическими животными того же размера. Крокодилу требуется от одной десятой до одной пятой части пищи, необходимой льву того же веса, и он может прожить без еды полгода.
Более низкие потребности в пище и адаптивный метаболизм позволяют рептилиям доминировать в животной жизни в регионах, где доступность пищи слишком мала для поддержания крупных млекопитающих и птиц [88] [89].
Принято считать, что рептилии не могут производить устойчиво высокий уровень энергии, необходимый для погони на большие расстояния или полёта. Более высокая энергетическая ёмкость могла быть ответственна за эволюцию теплокровности у птиц и млекопитающих. Однако исследование корреляции между активной ёмкостью и термофизиологией показывает слабую связь.
Большинство современных рептилий — плотоядные животные со стратегией кормления «сиди и жди»; неясно, являются ли рептилии хладнокровными из-за их экологии.
Энергетические исследования некоторых рептилий показали, что их активные способности равны или превосходят возможности теплокровных животных аналогичного размера [90] [91].
Вследствие разнообразия доступной пищи [ прояснить ] пищеварительный тракт у пресмыкающихся гораздо более дифференцирован, чем у земноводных животных. Пищеварительная система начинается ротовым отверстием, ограниченным челюстями с обычно коническими, одинаковыми, прочно прирастающими зубами гомодонтная система, хотя у некоторых вымерших форм была гетеродонтная.
У рептилий, за исключением крокодилов и большинства черепах, есть гибкие неподвижные губы, плотно закрывающие рот и зубы в состоянии покоя. Зубы пресмыкающихся не предназначены для жевания — они служат для захвата и удержания пищи, реже — для её разделения или отрывания удобных по размеру кусков, у видов, питающихся моллюсками и насекомыми, зубы приспособлены для дробления жёстких покровов этих животных. Пища практически всегда проглатывается целиком, как животная, так и растительная.
Язык свободный, впереди мускулистый, подвижный, к концу утончается и раздваивается. У некоторых видов язык клейкий, и служит для ловли добычи. Язык служит для проглатывания пищи, но виды с тонким и длинным языком проталкивают пищу в глотку толчками головы ящерицы или движением челюстей змеи Ротовая полость отграничена от глотки развивающимся вторичным костным нёбом у ящериц и черепах, полностью развитым костным небом у крокодилов, но у змей и варанов вторичное костное небо отсутствует.
Многоклеточные слюнные железы содержат пищеварительные ферменты. Глотка переходит в узкий, но очень растяжимый пищевод , далее в мускулистый желудок и кишечник. Желудок имеет толстые мышечные стенки. На границе между тонкой и толстой кишкой находится слепая кишка , которой нет у амфибий. Большая печень рептилий имеет жёлчный пузырь. Поджелудочная железа в виде длинного плотного тела лежит в петле двенадцатиперстной кишки.
Кишечник заканчивается клоакой.
Выделительная система пресмыкающихся представлена почками , мочеточниками и мочевым пузырём. Почки пресмыкающихся существенно отличаются от почек рыб и амфибий, которым приходится решать задачу избавления от постоянного избытка воды в организме. Вместо туловищных почек амфибий мезонефрос , почки рептилий метанефрос располагаются в тазовой области с брюшной стороны клоаки и по её бокам.
Почки соединяются с клоакой через мочеточники. Тонкостенный стебельчатый мочевой пузырь соединяется с клоакой тонкой шейкой на её брюшной стороне. У некоторых пресмыкающихся мочевой пузырь недоразвит крокодилы, змеи, некоторые ящерицы. У наземных рептилий в качестве конечного продукта азотистого обмена выступает слабо растворимая в воде мочевая кислота , выделяемая вместе с калом через клоаку, что позволяет более эффективно сохранять влагу. Половая система самцов состоит из пары семенников , которые расположены по бокам поясничного отдела позвоночника.
От каждого семенника отходит семенной канал , который впадает в вольфов канал. С появлением тазовой почки у пресмыкающихся вольфов канал у самцов выступает лишь как семяпровод и полностью отсутствует у самок. Вольфов канал открывается в клоаку, образуя семенной пузырёк. Половая система самок представлена яичниками , которые подвешены на брыжейке к спинной стороне полости тела по бокам позвоночника. Яйцеводы мюллеровы каналы также подвешены на брыжейке. В переднюю часть полости тела яйцеводы открываются щелевидными отверстиями — воронками.
Нижний конец яйцеводов открывается в нижний отдел клоаки на её спинной стороне. Оплодотворение внутреннее. Развитие зародыша происходит в яйце с кожистой или известковой оболочкой, наряду с этим встречается яйцеживорождение и реже истинное живорождение.
У пресмыкающихся прямое постэмбриональное развитие. Для многих представителей характерна забота о потомстве, в частности самки крокодилов переносят потомство от места кладки к водоёмам в ротовой полости, хотя в некоторых случаях могут съесть детёныша. Большинство пресмыкающихся относится к плотоядным животным включая насекомоядность и поедание падали, истинное хищничество среди современных рептилий свойственно только крокодилам , ряду ящериц и большинству змей.
Для некоторых например, агамы , игуаны характерно смешанное питание. Иногда встречаются и почти исключительно травоядные пресмыкающиеся сухопутные черепахи. Примитивные группы древних амниот в том числе первые рептилии изначально передвигались на четырёх широко расставленных в стороны конечностях, которые при локомоции раздвигались медиалатерально и не были способны постоянно поддерживать туловище над землёй, отсюда — «пресмыкание», волочение живота по земле, давшее русскоязычное название всей этой группе, что соответствовало характерному для них низкому уровню интенсивности обмена веществ и сравнительно малоподвижному образу жизни, связанному с чередованием коротких вспышек активности и длительного отдыха лёжа на животе.
Современные пресмыкающиеся в основном сохраняют подобный, базальный в эволюционном плане, характер локомоции, некоторое исключение составляют только крокодилы, у которых конечности при беге могут занимать положение почти под туловищем.
Причём у некоторых вымерших групп современного отряда крокодилов, которые вели исключительно сухопутный образ жизни, такое положение конечностей было габитуальным, то есть поддерживалось большую часть времени — в Австралии такие наземные крокодилы например, Квинкана обитали ещё 40 тыс. У варанов конечности полувыпрямлены, что связано с активным перемещением, и при беге туловище находится высоко над землёй. Змеи и безногие ящерицы так же является исключением, так как отсутствие конечностей и совершенно иной способ локомоции ползание , чем у рептилий с конечностями, обеспечивают быстрое перемещение большинству змей по любой поверхности.
Большинство рептилий неплохо плавают за счёт «рыбообразного» изгибания тела в поперечной плоскости. У некоторых групп приспособление к водной среде достигло более высокой степени, причём в процессе эволюции у них вырабатывались различные типы локомоции.
Так, ихтиозавры , мозазавры и, в меньшей степени, крокодилы сохранили характер плавания за счёт поперечных изгибов туловища. С другой стороны, у морских черепах и вымерших плезиозавров выработался новый механизм движения в воде, за счёт взмахов конечностей, преобразовавшихся в ласты. Многие эти группы практически полностью утратили способность перемещаться на суше. Несколько родов пресмыкающихся способны к планирующему полёту — например летучие драконы агамы рода Draco и украшенные древесные змеи.
Активно летающие рептилии известны только из ископаемых остатков см. В древности развитие некоторых продвинутых групп рептилий шло в иных направлениях — их конечности стали перемещаться непосредственно под туловище, занимая более габитуально вертикальное выпрямленное положение, что соответствовало более подвижному образу жизни, связанному с быстрым бегом и активным преследованием добычи. У предков архозавров лобковая и седалищная кости начали удлиняться, передние ноги — укорачиваться, а тяжёлый длинный хвост, практически лишившийся подвижности из-за срастания тазовых костей, превратился в балансир, уравновешивающий массу вытянутого туловища.
Задние конечности при ходьбе стали перемещаться в парасагиттальной плоскости, голеностопный сустав получил возможность пружинить при ходьбе, что способствовало более экономному расходу сил, а опора переместилась на средний третий палец.
Это позволило многим группам текодонтов перейти к факультативной, или даже габитуальной, бипедальности, подобно современным бегающим птицам, специализировавшись на беге на двух ногах. Данные изменения должны были сопровождаться значительным повышением интенсивности обмена веществ, поскольку постоянное поддержание тела над землёй и длительная локомоторная активность требовали намного больших энергозатрат по сравнению с «пресмыканием».
Однако проследить подобные изменения на палеонтологическом материале сложно. Достоверно известно, однако, что потомки данной группы рептилий — птицы — имеют интенсивный обмен веществ и эндотермную систему терморегуляции теплокровность. Ранние динозавры , как ящеротазовые , так и птицетазовые — были активными хищниками, как минимум часть времени передвигавшимися на задних конечностях, что способствовало их быстрому прогрессу, особенно после вымирания большинства синапсидов на рубеже пермского и триасового периода.
В юрском периоде почти все более-менее крупные наземные животные свыше метра в длину были динозаврами. Постепенно часть динозавров переходит на растительную пищу, занимая освободившиеся экологические ниши.
Большинство растительноядных видов вновь возвращается к передвижению на четырёх конечностях, эволюция же хищников продолжает идти по «двуногому» пути например, у тираннозаврид и абелизаврид передние конечности оказались практически редуцированны, а у манирапторов — напротив, развились в специализированную хватательную форму.
В течение млн лет, вплоть до конца мелового периода, основными наземными хищными позвоночными были двуногие динозавры, некоторые формы которых по большинству признаков неотличимы от птиц. Как видно, исторически для рептилий было свойственно главным образом именно активное передвижение на двух или четырёх выпрямленных конечностях, именно такие бегающие формы занимали господствующее положение в данном классе парафилетической группе животных на протяжении большей части его истории.
Двуногие хищные формы изредка возникали у птиц — например фороракосы. В настоящее время подобные формы среди рептилий не возникают, поскольку экологические ниши, связанные с активным преследованием добычи, ныне заняты млекопитающими, и организм современных рептилий требует значительной перестройки для их освоения.
Некоторое исключение составляет насекомоядная плащеносная ящерица , и некоторые другие агамовые, способные к бегу на двух ногах при сохранении расположения конечностей по бокам от туловища. Такая особенность позволяет им обходить ограничение Карриера невозможность бежать на четырёх ногах и дышать одновременно и активно преследовать добычу или спасаться от хищников.
Ящерица шлемоносный василиск во время бипедальной локомоции способна также бегать по поверхности воды. Тегу и вараны также научились обходить ограничение Карриера, но за счёт модификации дыхательной системы, а не подъёма на задние конечности.
Большинство рептилий не имеет настоящего голосового аппарата и производят лишь самые примитивные звуки вроде шипения или свиста.
Крокодилы рычат, издают лающие и рокочущие звуки, детёныши квакают. Черепахи в некоторых случаях испуг, брачный сезон, боль могут издавать писк или стон. Представители гекконовых ящериц издают самые разные звуки — писк, визг, щебет, щелчки, крики [92].
Значение пресмыкающихся для человека сравнительно невелико. Кожа крокодилов, крупных змей и ящериц применяется в кожевенной промышленности для изготовления чемоданов, ремней, обуви и т. Мясо и яйца многих черепах употребляются в пищу. Также могут употребляться в пищу некоторые ящерицы и змеи. Змеиный яд находит применение в медицине.
Многие змеи полезны истреблением грызунов, а ящерицы — насекомых. Некоторые виды пресмыкающихся держат в качестве домашних животных. Аллигаторы, кайманы и некоторые крокодилы иногда разводятся на специальных фермах, для получения мяса, яиц и кожи.
Большую опасность для людей представляют ядовитые змеи, особенно в тропических странах. Многие крокодилы опасны для человека, наносят значительный ущерб животноводству. Пресноводные черепахи вредят рыбному промыслу.
Крупные вараны, питоны и удавы так же могут быть опасны, будучи способными травмировать или даже убить человека. В году был расшифрован геном пресмыкающегося — ящерицы анолис каролинский [93].
Таким образом, это пресмыкающееся вошло в круг модельных объектов генетики. Материал из Википедии — свободной энциклопедии. Эта статья о парафилетической группе; о кладе см. Запросы «Рептилия» и «Рептилии» перенаправляются сюда; см. Без ранга:. Основная статья: Происхождение рептилий. Оригинальный текст англ. Mammals are a clade, and therefore the cladists are happy to acknowledge the traditional taxon Mammalia; and birds, too, are a clade, universally ascribed to the formal taxon Aves.
Mammalia and Aves are, in fact, subclades within the grand clade of the Amniota. But the traditional class Reptilia is not a clade. It is just a section of the clade Amniota: the section that is left after the Mammalia and Aves have been hived off. It cannot be defined by synapomorphies, as is the proper way. Instead, it is defined by a combination of the features it has and the features it lacks: reptiles are the amniotes that lack fur or feathers.
Pareiasaurus serridens Procolophonomorpha. Platypterygius kiprijanoffi Ichthyosauria. Лепидозавры Lepidosauria. По часовой стрелке, начиная сверху слева: гаттерия , чёрная мамба , обыкновенная игуана , Smaug breyeri и сетчатый питон. Черепахи Testudines. Сверху: краснобрюхая короткошейная черепаха , индийская лопастная черепаха ; Снизу: галапагосская черепаха , бисса. Архозавриформы Archosauriformes. В разделе не хватает ссылок на источники см. Информация должна быть проверяема , иначе она может быть удалена.
Вы можете отредактировать статью, добавив ссылки на авторитетные источники в виде сносок. A reevaluation of early amniote phylogeny англ. Архивировано 8 июня года. Observations of the structure of the Early Permian reptile Stereosternum tumidum Cope англ. The phylogenetic definition of Reptilia англ.
Архивировано 3 ноября года. Reptile Database. Дата обращения: 17 ноября Архивировано 14 ноября года. Архивировано 2 июня года. Дата обращения: 10 сентября Архивировано 4 июня года.
Архивировано 16 июля года. The First Age of Reptiles? Архивировано 25 сентября года. Архивировано 28 мая года. Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed англ. Архивировано 3 декабря года. Neto, and Pedro L. Архивировано 11 сентября года. Tuinen, E. Vertebrate palaeontology.
Rieppel, M. Дата обращения: 11 сентября Katsu, E. Braun, L. Guillette Jr, T. Архивировано 12 ноября года. Ichthyosaurian relationships illuminated by new primitive skeletons from Japan англ. Архивировано 19 октября года. Evans, Jozef Klembara. Hansen, C. Josh Donlan, Christine J.
Griffiths, Karl J. Ecological history and latent conservation potential: large and giant tortoises as a model for taxon substitutions англ.
Архивировано 8 августа года. Longrich, Bhart-Anjan S. Bhullar and Jacques A. Архивировано 16 ноября года. Архивировано 3 июня года. Reeder, Ted M. Townsend, Daniel G. Mulcahy, Brice P. Noonan, Perry L. Wood, Jr. Sites, Jr. Архивировано 4 февраля года. Reading the world : encyclopedic writing in the scholastic age. Caroli Linnaei Systema naturae per regna tria naturae :secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis.
Laurentii Salvii, Архивировано 10 октября года. Архивировано 28 июня года. Архивировано 17 августа года. Архивировано 10 сентября года. Series B, Biological Sciences. Архивировано 1 января года. The variety of life : a survey and a celebration of all the creatures that have ever lived. Vertebrate Paleontology. Tsuji, J. Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade англ.
Архивировано 3 августа года. Архивировано 22 февраля года. Fossil Reptiles of Great Britain англ. Vertebrate palaeontology ru en англ. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae англ. Архивировано 18 сентября года. В прошлом палеонтологи считали, что теплокровность и связанный с ней высокий уровень активности в любое время суток были одной из самых древних и важных особенностей млекопитающих, благодаря этому эти животные занимали те экологические ниши, на которые не могли претендовать холоднокровные рептилии.
Ньюэм отмечает, что это не обязательно говорит в пользу того, что морганукодоны и куэнотерии не были теплокровными. Но это говорит о том, что древние родичи современных млекопитающих значительно уступали им в уровне активности.
Как и когда древние млекопитающие стали более похожими на современных, ученым еще предстоит выяснить. В черных дырах могут быть вселенные. Рассказываем о новом открытии. Из микропузырьков графена сделали идеальные линзы. Закрыть меню. Поиск по сайту. Последние материалы Быстрая связь с Марсом возможна с помощью квантовых технологий, считают ученые В Японии планируют использовать ракеты, работающие на топливе из коровьего навоза. О проекте Реклама Контакты Рассылка.
Подписка на дайджест Ввод e-mail для подписки Email Подписаться. Даю согласие на обработку моих персональных данных в соответствие с Политикой Конфиденциальности и Пользовательским соглашением.
Выключить push-уведомления. Политика обработки персональных данных. Наука 12 октября Первые млекопитающие были похожи на рептилий по скорости обмена веществ.